LES GENES
LEUR TRADUCTION
A - Le dictionnaire
L'ADN nucléaire contient le code génétique de la cellule. Ce code compte quatre lettres (A, G, C et T ou U), (T) dans l'ADN et (U) dans l'ARN. Ces quatre lettres groupées par deux forment 16 paires (4²), nombre insuffisant pour coder les 20 acides aminés. Aussi, la nature a-t-elle ajouté une troisième lettre. La correspondance de ces triplets ou codons avec les 20 acides aminés est donnée sur le Tableau n° 134 ci-dessous. On y constate que la troisième lettre est quelque peu aléatoire.
Nous y avons mentionné les codons actuellement utilisés et ceux dits primitifs.
Les ACIDES AMINES ou AMINOACYLS
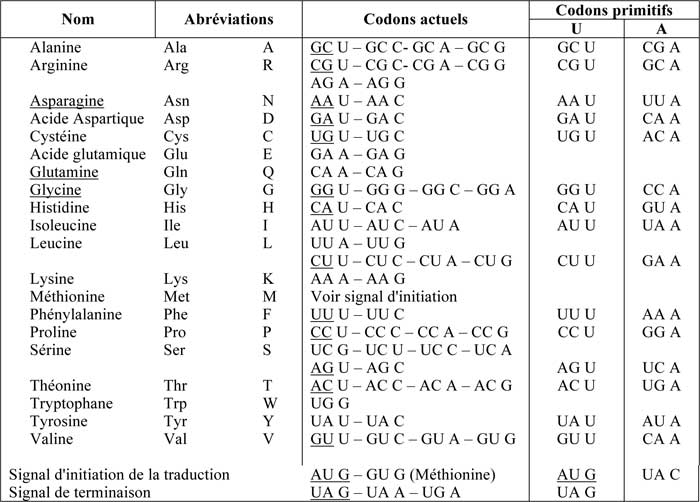
U : sont les codons supposés primitifs.
A : leur anti-codon
Il faut remarquer que certains biologistes présentent les anti-codons dans un sens de lecture inverse ; par exemple, l'Alanine codon : GCU, anticodon CGA, lequel est inversé : AGC
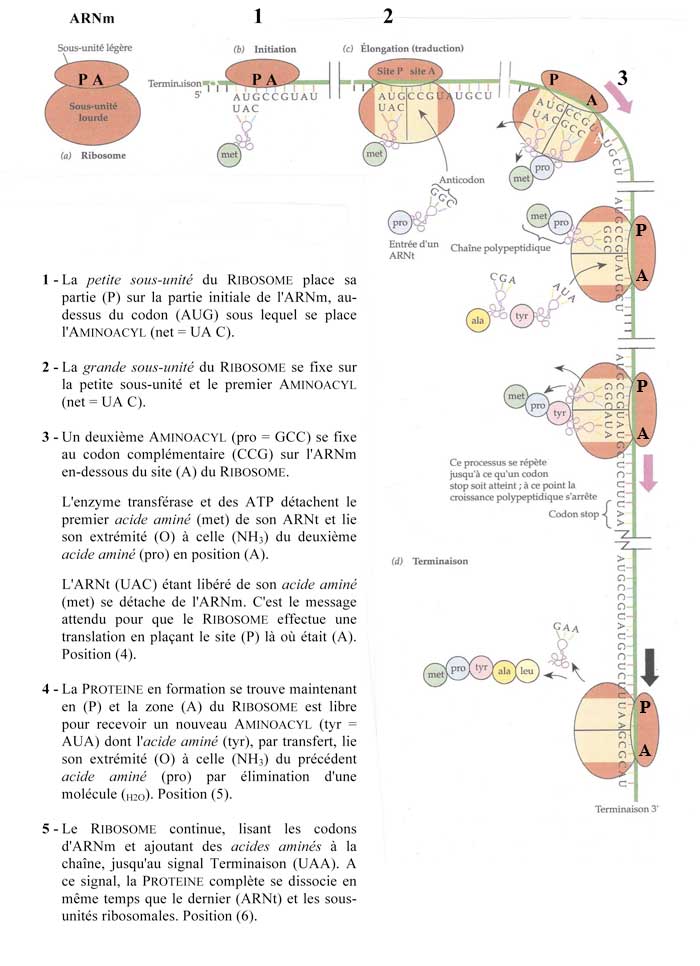
B - La traduction consiste à assembler les acides aminés pour en faire des PROTEINES à partir des informations contenues dans l'ARNm mature à l'aide des ARNt (transfert) et des RIBOSOMES selon le processus de la figure suivante.
C - L'adressage des protéines
Les protéines des eucaryotes doivent, après leur synthèse, atteindre leur cible finale, le cytoplasme, la membrane ou un organite quelconque. Il peut y avoir des membranes à traverser. En fait, c'est le cas pour la plupart des protéines, seules les protéines cytoplasmiques sont produites directement sur leur lieu d'utilisation, mais même pour elle, leur localisation n'est pas due au seul hasard, autrement l'organisation de la cellule serait passablement perturbée.
On peut imaginer un système de translocation qui permettrait aux protéines de passer les membranes qui les empêchent d'atteindre leur cible. Une fois les membranes traversées, il n'y a plus de problème, la localisation dans le compartiment membranaire ou liquidien de l'organite dépend de l'hydrophobicité de la protéine. De tels systèmes de translocation existent en effet. On les trouve sur les membranes des principaux organites : noyau, réticulum, mitochondries, plastes.
Le phénomène qui permet aux protéines d'atteindre leur cible finale est l'adressage. Il fonctionne de la façon suivante : toute protéine spécifique d'un organite débute par une séquence signal ou peptide, signal spécifique de la cible. Il y peut y en avoir plusieurs à la suite pour affiner la destination. Les protéines de la matrice des mitochondries, par exemple, ont deux membranes à traverser et donc deux translocations à subir. Pour le réticulum, c'est encore plus complexe, une protéine exportée dans sa lumière pourra soit être intrinsèque au réticulum, soit au golgi, soit intégrée aux vésicules d'exocytoses, soit aux lysosomes, soit à la membrane, chaque cas correspond à un peptide signal qui suit celui du signal de localisation réticulaire plus général. Dès que le peptide signal est synthétisé, il est reconnu par le système de translocation qui fait traverser le pore à la chaîne protéique. Tout de suite après la traversée du pore, il est excisé par un complexe enzymatique. A partir de là, la synthèse se poursuit à la chaîne étant injectée dans le pore au fur et à mesure de son élongation.
Il est finalement amusant de constater que la cellule, pour acheminer une protéine à sa destination finale, utilise la même technique que l'homme pour acheminer son courrier : elle marque l'adresse dessus.
D - Quelques remarques
Plusieurs RIBOSOMES se déplacent simultanément sur un même ARNm pour traduire la même protéine, tels des ouvriers devant un tapis roulant garni de pièces détachées qui assembleraient le même appareil.
Lorsqu'une protéine est fabriquée, elle n'est qu'un long polymère qui ne tarde pas à se mettre spontanément en mouvement, se replie, vrille une extrémité, recroqueville l'un de ses fragments sur lui-même jusqu'à former un édifice tridimentionnel dont la forme détermine sa fonction biologique.
La séquence en acides aminés d'une protéine constitue sa structure "primaire". Les brins
repliés, les feuillets, les hélices,… sont ses structures "secondaires". L'assemblage de tous
les éléments secondaires correspond à la structure tertiaire
.
Les protéines destinées à l'usage externe de la cellule sont acheminées dans des vésicules qui fusionnent avec la paroi de l'appareil de GOLGI où elles subissent la glycocylation. La protéine se retrouve liée à une molécule qui la transporte jusqu'à la membrane cellulaire pour être expulsée à l'extérieur.
L'appareil de GOLGI sert de réservoir dans lequel les protéines synthétisées sont conservées, modifiées et empaquetées pour être délivrées à l'extérieur de la cellule ou dans d'autres organites.
L'ARNm mature du CYTOPLASME peut être relativement stable pendant quelques heures, voire quelques jours, mais de toute façon, il se dégrade.
L'ARNm mature est aussi à l'origine des ARN polymérase, maturase, etc.

